
Fig. 1. Development of "determinate axes" (arrows) into new axes near the thallus base in Balliella cladoderma (Athanasiadis 1996, p. 41).
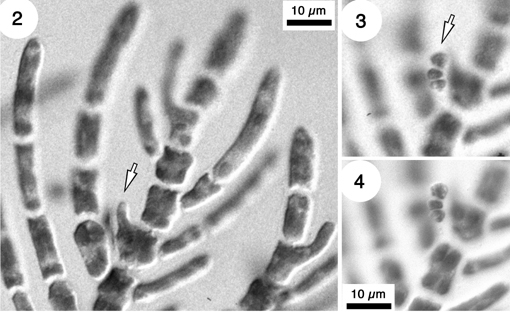
Figs 2-4. Development of a 3-celled carpogonial branch (arrow in fig. 3, 4) on a laterally dividing axial cell (arrow in fig. 2) in Balliella cladoderma (Athanasiadis 1996, p. 41).

Figs 5-9. Development of gland cells in Balliella cladoderma. Gland cells are cut off from axial cells (white arrow in fig. 5) and are transferred to periaxial cells either abaxially (black arrow in fig. 5, fig. 6, arrow in fig. 7) or adaxially (fig. 8). The final position of gland cells is either abaxial (black arrow in fig. 9) or adaxial (white arrows in fig. 9).
Fig. 10. Gland cells in Scageliopsis strongylekystis where the gland cell touches only the mother cell (Athanasiadis 1996, p. 180).
Fig. 11. Gland cells in Antithamnion cruciatum where the gland cells lie mainly on the contiguous cells touching slightly the mother cell.

Fig. 12. Post-fertilization in Pterothamnion squarrulosum where the auxiliary cell has cut off an apical cell to receive the zygote (Athanasiadis & Kraft 1994).
Fig. 13. Post-fertilization in Antithamnionella spirographidis where the carpogonium has developed an extension tube to transfer the zygote (Athanasiadis 1996, p. 121).
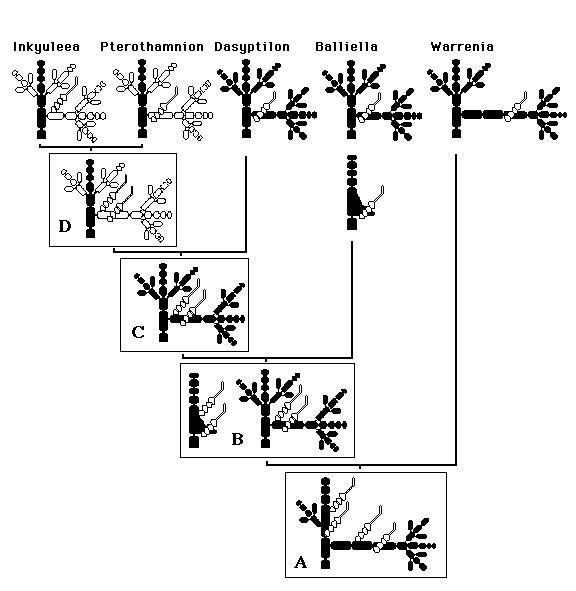
Fig. 14. Hypothetical ontogeny of adaxial and abaxial carpogonial branches (c.bs) in the Ceramiales.
A. The ancestral type where c.bs developed on cells of main and lateral filaments of unlimited growth (Athanasiadis 1996, p. 36).
B. The ancestral type of Balliella, where c.bs developed on axial cells and were transferred to periaxial cells either adaxially or abaxially (only abaxial c.bs have persisted in Balliella).
C. The ancestral type of the Callithamnioideae, where c.bs developed directly on periaxial cells either abaxially or adaxially (only abaxial c.bs have persisted in the Callithamnioideae).
D. The ancestral type of the Ceramioideae that gave rise to abaxial c.bs (as generally occurs in the subfamily) and adaxial c.bs (as occurs in Inkyuleea).
Frames indicate hypothetical types. Shaded filaments are of unlimited growth and unshaded filaments are of limited growth.

Fig. 15. Cladograms reflecting relationships between genera, tribes and families of the Ceramiales. Modified after:
A. Morphological data (Athanasiadis 1996).
B. SSU rRNA sequences (Saunders et al. 1996).
C. RbcL sequences (Jong et al. 1998).
D. SSU rRNA sequences (Choi et al. 2000), and
E. LSU rDNA and SSU rDNA sequences (Harper & Saunders 2001).

Fig. 16. Relationships between six primitive genera of Ceramiaceae as indicated by the most parsimonious distribution of 12 characters (Table I) or character states (within 7 characters; Table II). Periaxial vs. axial position of carpogonial branches refers to the final position and not to the place of origin. Loss of gland cells (in Inkyuleea and in the ancestor of the Callithamnioideae) is accepted as a parallelism (instead of a reduction for the Ceramiaceae and a reversal for Pterothamnion).
