Common Name: SEDGE FAMILY Habit: Annual, perennial herb, often rhizomed or stoloned, often of wet open places; roots fibrous; monoecious, dioecious, or flowers bisexual. Stem: generally 3-sided, generally solid. Leaf: generally 3-ranked; base sheathing, sheath generally closed, ligule generally 0; blade (0 or) linear, parallel-veined. Inflorescence: spikelets generally arranged in head-, spike-, raceme-, or panicle-like inflorescences; flower generally sessile in axil of flower bract, enclosed in a sac-like structure (perigynium) or generally not. Flower: unisexual or bisexual, small, generally wind-pollinated; perianth 0 or generally bristle like; stamens generally 3, anthers attached at base, 4 chambered; ovary superior, chamber 1, ovule 1, style 2--3(4)-branched. Fruit: achene, 2--3 sided. Genera In Family: +- 100 genera, 5000 species: especially temperate. Note: Difficult; taxa differ in technical characters of inflorescence, fruit. In Carex and Kobresia, what appear to be individual pistillate flowers in fact are highly reduced inflorescences (whether or not the same applies to staminate flowers is still under debate). In some other works (e.g., FNANM) these are called spikelets, and they are treated as being arranged in spikes. Here and in TJM (1993), what appear to be individual pistillate flowers are called pistillate flowers in Carex (and they are treated as being arranged in spikelets), but spikelets in Kobresia (and they are treated as being arranged into spikes). Though internally inconsistent, the approach here is consistent with traditional usage, and reflects a preference for character states that may be determined in the field. Molecular, morphological, and embryological evidence indicates that Eriophorum crinigerum is to be segregated to a new genus, as Calliscirpus criniger (A. Gray) C.N. Gilmour et al., along with a second, newly described species, Calliscirpus brachythrix C.N. Gilmour et al. (Gilmour et al. 2013); key to genera modified by Peter W. Ball to include Calliscirpus. eFlora Treatment Author: S. Galen Smith, except as noted Scientific Editor: S. Galen Smith, Thomas J. Rosatti, Bruce G. Baldwin.
Common Name: SEDGE Habit: Perennial herb, cespitose to loosely cespitose to rhizomed with internodes > 1 cm; generally monoecious. Stem: generally sharp-3-angled, generally solid. Leaf: 3-ranked, generally glabrous except generally scabrous on midrib, margin; sheath closed, back (blade side of stem) green, ribbed, front (non-blade side of stem) generally thin, translucent, sometimes cross-wrinkled or flat, forming generally U-shaped mouth at top, sometimes extending above blade as a fragile sleeve-like "contraligule" (especially Groups 7, 11), sometimes disintegrating to a ladder- or lattice-like network or fringe of veins ("leaf sheath fronts fibrous"). Inflorescence: spikelets generally several to many, in spike, raceme, panicle, or head-like arrangement, each 1--many-flowered, generally unisexual, or bisexual, then staminate flowers distal to pistillate ("staminate/pistillate"), pistillate distal to staminate ("pistillate/staminate"), or otherwise, generally subtended by spikelet bract, lowest subtended by inflorescence bract, occasionally some additional pistillate spikelets on lateral shoots from basal nodes ("basal spikelets"); flowers subtended by flowering bract ("scale" in other literature, especially for pistillate). Flower: unisexual; perianth 0. Staminate Flower: stamens generally 3. Pistillate Flower: enclosed by sac-like structure (perigynium, abbreviated to "peri" here), occasionally next to bristle-like axis; style 1, stigmas 2--3(4), exserted. Fruit: 2--3(4)-sided, enclosed in peri, stalked or not, style base generally not persistent; peri body 2--3(4)-sided or round, often with marginal ribs, some with additional veins, papillate or not (determined at 20×), abruptly narrowed at base into stalk or not; peri beak abaxial flap (suture) prominent or generally inconspicuous or 0, tip open, often notched. Etymology: (Latin: cutter, from sharp leaf, stem edges) Note: Difficult because of many species, morphologic and genetic variation, minute key characters. Peri around fully mature fruit needed for identification (long-persistent peri often atypical). Many herbarium specimens have immature peri, which lead to misidentification. 2-styled plants with peri +- flat adaxially, curved abaxially are planoconvex; peri curved +- equally on both surfaces are biconvex. Peri walls said to be translucent are easily punctured and/or do not completely conceal fruit within. Peri beaks generally measured from point of inflection, where peri margin changes from convex to concave, to its tip, but in a few taxa it is measured from fruit top to beak tip ("measured from fruit top" for those taxa). Peri (and fruit) shapes including beak; peri (and fruit) "body" excludes beak. Mid to late season shoots often atypical in shape, color of inflorescence, bracts, peri. Number of peri given is per spikelet. Actual hybrids probably less frequent than reports of hybrids. Carex pityophila Mack, native to southern Rocky Mountains, reported from SnBr, but is distinct; plants from SnBr warrant a new species. In TJM (1993), Carex cephalophora Willd. misappl. to plants belonging instead to Carex mesochorea Mack. (Group 9), native to eastern United States, collected in SCo (Los Angeles Co.) in 1929 and in ScV (Butte Co.) in 2010. Carex molesta Mack. ex Bright (Groups 11A,G), native to eastern United States, an historical urban weed, Carex leavenworthii Dewey an urban weed. Carex cyrtostachya, Carex orestera, and Carex xerophila described since TJM2. Plants called Carex albonigra_in TJM2 are Carex orestera; Carex albonigra not in California. Unabridged Note:Carex molesta Mack. ex Bright (Groups 11A,G), native to eastern United States, collected once from a parking space in SCo (Santa Barbara Co.) in 1958, making it an historical urban weed. eFlora Treatment Author: Peter F. Zika, Andrew L. Hipp & Joy Mastrogiuseppe Reference: Wilson et al. 2007 J Bot Res Inst Texas 1:69--77; Zika et al. 1998 Madroño 45:261--270 Unabridged Reference: Ball, P. W. and A.A. Reznicek. 2002. Carex, In: Flora of North America Editorial Comittee, Eds. Flora of North America North of Mexico. Volume 23, Magnoliophyta: Commelinidae (in part): Cyperaceae. Oxford University Press, New York. pp. 254--572; Dean, E., F. Hrusa, G. Leppig, A. Sanders, and B. Ertter. 2008. Catalogue of nonnative vascular plants occuring spontaneously in California beyond those addressed in the Jepson Manual - part II. Madroño 55: 93--112; Goldman, D. 2008. Noteworthy collections, California. Carex longii. Madroño 55: 89--90; Hipp, A. L., A.A. Reznicek, P.E. Rothrock and J.A. Weber. 2006. Phylogeny and classification of Carex section Ovales (Cyperaceae). International Journal of Plant Sciences 167: 1029--1048; Hipp, A. L., P.E. Rothrock, A.A. Reznicek, and P.E. Berry. 2007. Chromosome number changes associated with speciation in sedges: A phylogenetic study in Carex section Ovales (Cyperaceae) using AFLP data. Aliso 23: 193--203; Hipp, A. L., P.E. Rothrock and E.H. Roalson. 2008. The evolution of chromosome arrangements in Carex (Cyperaceae). The Botanical Review 75: 96--109; Janeway, L. P. 1992. Cyperaceae of Butte County, California. Part 1: Carex. Studies from the herbarium (Number 9), California State University, Chico; Mackenzie, K. K. 1917. Notes on Carex-XI. Californian representatives of the Ovales. Bulletin of the Torrey Botanical Club 43: 601--620; Mackenzie, K. K. 1922. A monograph of the California species of the genus Carex. Erythea 8: 7--95; Murray, D. F. 1969. Taxonomy of Carex sect. Atratae (Cyperaceae) in the southern Rocky Mountains. Brittonia 21: 55--76; Naczi, R. F., C.T. Bryson, and T.S. Cochrane. 2002. Seven new species and one new combination in Carex (Cyperaceae) from North America. Novon 12: 508--532; Reznicek, A. A. 1993. Revision of Carex section Ovales (Cyperaceae) in Mexico. Contributions form the University of Michigan Herbarium 19: 97--136; Reznicek, A. A. and P.W. Ball. 1980. The taxonomy of Carex section Stellulatae in North America north of Mexico. Contributions from the University of Michigan Herbarium 14: 153--203; Rothrock, P. E., and A.A. Reznicek. 2001. The taxonomy of the Carex bicknellii group (Cyperaceae) and new species for central North America. Novon 11: 205--228; Standley, L. A. 1985. Systematics of the Acutae group of Carex (Cyperaceae) in the Pacific Northwest. Syst Bot Monographs 7: 1--106; Taylor, D. W., and J. Mastrogiuseppe. 1999. Carex tiogana (Cyperaceae), a new sedge from the Sierra Nevada, California. Novon 9: 120--123; Wilson, B. L., R.E. Brainerd, L.P. Janeway, K. Kuykendall, D. Lytjen, B. Newhouse, N. Otting, S. Meyers, and P. Zika. 2007. Description of Carex klamathensis (Cyperaceae), a rare sedge of the Klamath Region of Oregon and California, U.S.A.J. Bot. Res. Inst. Texas 1: 69--77; Wilson, B. L., R. Brainerd, D. Lytjen. B. Newhouse, and N. Otting. 2008. Field Guide to the Sedges of the Pacific Northwest. Oregon State University Press, Corvallis, OR; Zika, P. F., K. Kuykendall, and B. Wilson. 1998. Carex serpenticola (Cyperaceae), a new species from the Klamath Mountains of Oregon and California. Madroño 45: 261--270; Zika, P. F. 2012. Carex orestera (Cyperaceae), a new sedge from the mountains of California. Novon 22: 118--124; Zika, P. F. and B. L. Wilson. 2012. Carex albida (Cyperaceae), and its relationship to Carex lemmonii. Madroño 59: 171--180; Zika, P. F., L. P. Janeway, B. L. Wilson, and L. Ahart. 2013. Carex crytostachya (Cyperaceae), a new species of sedge endemic to the Sierra Nevada of California. Journal of the Botanical Research Institute of Texas 7: 25--35; Zika, P. F., L. P. Janeway, and B. L. Wilson. 2014. Carex xerophila (Cyperaceae), a new sedge from the chaparral of northern California. Madroño 61: 299--307.
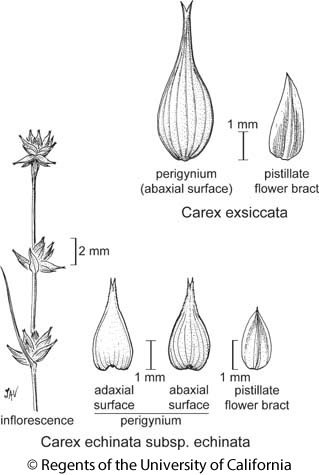
Stem: edges +- blunt. Inflorescence: spikelets 3--15.5 mm, outline +- jagged at arm's length, base of terminal narrow-tapered; inflorescence bract << inflorescence; staminate flower bracts of terminal spikelet appressed, internodes between them 2/3--3/4 their length; pistillate flower bract brown, white-margined, generally acute. Fruit: peri spreading to reflexed, generally 2.9--4.75 mm, widest +- at base, length 1.7--3.6 × width, tapered to beak, green to brown, lower wall filled with pithy tissue, beak generally 0.95--2 mm, 35--86% length of body, sparse-serrate, tip +- brown.
Carex echinata Murray subsp. echinata
NATIVE Leaf: blade generally 0.7--2.7 mm at widest. Inflorescence: open to dense, 0.7--7.8 cm. Fruit: 1.1--1.6 mm, 0.8--1.3 mm wide, +- round; peri generally 2.9--4 mm, adaxially generally unveined, with bulge of pithy tissue below fruit, body generally serrate. Chromosomes: 2n=50,52,58. Ecology: Wet places, especially sphagnum bogs; Elevation: < 3200 m. Bioregional Distribution: KR, NCoRO, CaRH, SNH, SnBr; Distribution Outside California: to British Columbia, eastern North America, Central America, Eurasia, Australia. Fruiting Time: Jun--Sep Synonyms: Carex angustior Mack.; Carex ormantha (Fernald) Mack. Jepson eFlora Author: Peter F. Zika, Andrew L. Hipp & Joy Mastrogiuseppe Reference: Wilson et al. 2007 J Bot Res Inst Texas 1:69--77; Zika et al. 1998 Madroño 45:261--270 Index of California Plant Names (ICPN; linked via the Jepson Online Interchange) Previous taxon: Carex echinata Next taxon: Carex echinata subsp. phyllomanica
Botanical illustration including Carex echinata subsp. echinata
Citation for this treatment: Peter F. Zika, Andrew L. Hipp & Joy Mastrogiuseppe 2015, Carex echinata subsp. echinata, in Jepson Flora Project (eds.) Jepson eFlora, Revision 3, https://ucjeps.berkeley.edu/eflora/eflora_display.php?tid=49674, accessed on April 24, 2024.
Citation for the whole project: Jepson Flora Project (eds.) 2024, Jepson eFlora, https://ucjeps.berkeley.edu/eflora/, accessed on April 24, 2024.
MAP CONTROLS 1. You can change the display of the base map layer control box in the upper right-hand corner.
2. County and Jepson Region polygons can be turned off and on using the check boxes.
(Note: any qualifiers in the taxon distribution description, such as 'northern', 'southern', 'adjacent' etc., are not reflected in the map above, and in some cases indication of a taxon in a subdivision is based on a single collection or author-verified occurence).
Data provided by the participants of the
Consortium of California Herbaria.
MAP LEGEND View all CCH records All markers link to CCH specimen records. The original determination is shown in the popup window.
Blue markers indicate specimens that map to one of the expected Jepson geographic subdivisions (see left map). Purple markers indicate specimens collected from a garden, greenhouse, or other non-wild location.
Yellow markers indicate records that may provide evidence for eFlora range revision or may have georeferencing or identification issues.
READ ABOUT YELLOW FLAGS
CCH collections by month
Duplicates counted once; synonyms included.
Species do not include records of infraspecific taxa, if there are more than 1 infraspecific taxon in CA.
Blue line denotes eFlora flowering time (fruiting time in some monocot genera).