Common Name: SEDGE FAMILY Habit: Annual, perennial herb, often rhizomed or stoloned, often of wet open places; roots fibrous; monoecious, dioecious, or flowers bisexual. Stem: generally 3-sided, generally solid. Leaf: generally 3-ranked; base sheathing, sheath generally closed, ligule generally 0; blade (0 or) linear, parallel-veined. Inflorescence: spikelets generally arranged in head-, spike-, raceme-, or panicle-like inflorescences; flower generally sessile in axil of flower bract, enclosed in a sac-like structure (perigynium) or generally not. Flower: unisexual or bisexual, small, generally wind-pollinated; perianth 0 or generally bristle like; stamens generally 3, anthers attached at base, 4 chambered; ovary superior, chamber 1, ovule 1, style 2--3(4)-branched. Fruit: achene, 2--3 sided. Genera In Family: +- 100 genera, 5000 species: especially temperate. Note: Difficult; taxa differ in technical characters of inflorescence, fruit. In Carex and Kobresia, what appear to be individual pistillate flowers in fact are highly reduced inflorescences (whether or not the same applies to staminate flowers is still under debate). In some other works (e.g., FNANM) these are called spikelets, and they are treated as being arranged in spikes. Here and in TJM (1993), what appear to be individual pistillate flowers are called pistillate flowers in Carex (and they are treated as being arranged in spikelets), but spikelets in Kobresia (and they are treated as being arranged into spikes). Though internally inconsistent, the approach here is consistent with traditional usage, and reflects a preference for character states that may be determined in the field. Molecular, morphological, and embryological evidence indicates that Eriophorum crinigerum is to be segregated to a new genus, as Calliscirpus criniger (A. Gray) C.N. Gilmour et al., along with a second, newly described species, Calliscirpus brachythrix C.N. Gilmour et al. (Gilmour et al. 2013); key to genera modified by Peter W. Ball to include Calliscirpus. eFlora Treatment Author: S. Galen Smith, except as noted Scientific Editor: S. Galen Smith, Thomas J. Rosatti, Bruce G. Baldwin.
Common Name: SPIKERUSH Habit: Annual, perennial herb, generally forming mats, glabrous, internal air cavities evident; caudex generally 0; rhizomes generally evident, long, scaly, bulb or tuber at tip generally 0. Stem: simple, generally erect, smooth, generally not hollow; tip generally not rooting. Leaf: 2, basal, blades 0 or tooth-like, <= 1 mm. Inflorescence: inflorescence bracts 0; spikelet terminal, 1, generally ovate, not +- flat [(+- flat)], generally not forming plantlets, flowers 3--100+; flower bracts spiraled [(2-ranked)], each with 1 flower in axil, generally ovate, generally brown, generally membranous, smooth, tip generally acute to obtuse, notch 0; basal flower bract generally encircling stem, generally < 1/2 spikelet, flower generally 0. Flower: bisexual; perianth parts reduced to bristles, 0--8, generally +- <= fruit, barbs generally recurved; stamens generally 3; style 1, thread-like, base enlarged, generally persistent on fruit as tubercle. Fruit: generally obovate, generally brown; tubercle (0 or) generally distinct, generally pyramidal. Etymology: (Greek heleios, dwelling in a marsh, and Charis, grace) Note:Eleocharis lanceolata Fernald, Eleocharis equisetoides Torr. not in California. eFlora Treatment Author: S. Galen Smith Reference: Smith et al. 2002 FNANM 23:60--120
Eleocharis geniculata (L.) Roem. & Schult.
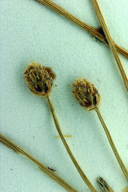
NATIVE Habit: Annual 5--45 cm, tufted; rhizome 0. Stem: 0.2--1 mm diam, cylindric. Leaf: distal sheath firm, persistent, tip acute. Inflorescence: spikelet 1--9 mm, 1--4 mm wide; flower bracts to 125, 0.8--3 mm, membranous to papery, rounded to acute. Flower: anthers +- 0.5 mm; stigmas 2. Fruit: 0.5--1.1 mm, 0.3--0.7 mm wide, 2-sided, smooth, black (brown when immature); perianth bristles (0)4--8, vestigial to generally = fruit (much exceeding tubercle), generally red-brown. Chromosomes: 2n=10. Ecology: Uncommon, very local in California. Wet, often brackish places; Elevation: < 1500 m. Bioregional Distribution: SCo/PR, SnBr, DSon; Distribution Outside California: to southern Ontario, Florida; Mexico, West Indies, South America, Asia, Africa, Pacific Islands. Flowering Time: Spring--winter Synonyms: Scirpus geniculatus L.; Eleocharis capitata R. Br.; Eleocharis caribaea (Rottb.) S.F. Blake; Eleocharis dispar E.J. Hill Jepson eFlora Author: S. Galen Smith Reference: Smith et al. 2002 FNANM 23:60--120 Index of California Plant Names (ICPN; linked via the Jepson Online Interchange) Previous taxon: Eleocharis flavescens var. flavescens Next taxon: Eleocharis macrostachya
Botanical illustration including Eleocharis geniculata
Citation for this treatment: S. Galen Smith 2012, Eleocharis geniculata, in Jepson Flora Project (eds.) Jepson eFlora, https://ucjeps.berkeley.edu/eflora/eflora_display.php?tid=23962, accessed on April 25, 2024.
Citation for the whole project: Jepson Flora Project (eds.) 2024, Jepson eFlora, https://ucjeps.berkeley.edu/eflora/, accessed on April 25, 2024.
MAP CONTROLS 1. You can change the display of the base map layer control box in the upper right-hand corner.
2. County and Jepson Region polygons can be turned off and on using the check boxes.
(Note: any qualifiers in the taxon distribution description, such as 'northern', 'southern', 'adjacent' etc., are not reflected in the map above, and in some cases indication of a taxon in a subdivision is based on a single collection or author-verified occurence).
Data provided by the participants of the
Consortium of California Herbaria.
MAP LEGEND View all CCH records All markers link to CCH specimen records. The original determination is shown in the popup window.
Blue markers indicate specimens that map to one of the expected Jepson geographic subdivisions (see left map). Purple markers indicate specimens collected from a garden, greenhouse, or other non-wild location.
Yellow markers indicate records that may provide evidence for eFlora range revision or may have georeferencing or identification issues.
READ ABOUT YELLOW FLAGS
CCH collections by month
Duplicates counted once; synonyms included.
Species do not include records of infraspecific taxa, if there are more than 1 infraspecific taxon in CA.
Blue line denotes eFlora flowering time (fruiting time in some monocot genera).